I'm calling this an article about genetics. It is such. But -- fair warning -- it is also, and most especially, an article about evolution, because genetics and evolution are inextricably linked. Obviously this is controversial, but I have to do my best to cover all aspects of textual criticism. And, if you study evolution, and then read this article, you will surely see why I decided I had to write this.
Much of this has been very difficult to write; I've already produced at least half a dozen different versions just of this disclaimer, doubtless spending vastly more time than it's worth. I've tried to figure out a way to finesse this -- to argue that the question of whether evolution is "real" is not very important -- but I don't think the argument can be sustained. You can be a good textual critic whether you believe in evolution or not -- but, after some months of study of the recent changes in the understanding of the theory of evolution, I am completely convinced that you will be a better one if you can learn from what we know about evolution. This apart from the fact that a correct understanding of evolution is vital in politics, because the implications of evolution require certain policies which politicians and the general population are often unwilling to admit. (There is nothing unique about that. The same goes at least double for thermodynamics. There are times when I think that the best way to succeed in American politics is to go into therapy to have all knowledge of science erased from one's brain.)
So let it be stated: I believe in evolution. I beleive it because all life on earth shares the same genetic code -- that is, all species use the same DNA encoding for amino acids, meaning that a gene from, say, corn, can be inserted into, say, an ape and still produce the same protein. This means that bacteria and viruses can (and do) share genes, meaning that evolution, even if it didn't happen in the past, will happen in the future. I believe in evolution because it's happening right now. Indeed, it can be made to happen in the lab. I believe in it because it explains so much that is bad about human behavior -- wars, murders, crime, false advertising. Indeed, one recent theory has it that the reason humans developed large brains is for purposes of manipulating and tricking each other. There are only two species in which males band together to make war on their neighbours. They are human beings -- and, it turns out, chimpanzees, our closest relatives. (And we thought they were our closest relatives even before this behavior was discovered.) I don't much like evolution, given how it seems to be working out. Much as I'd like to deny it, I can't see a way to avoid it.
Let me offer an analogy as to why I think it matters even if perhaps it isn't "true." If evolution did not create biodiversity, then the Creator created it to look as if evolution created biodiversity, by supplying fossils and such. This is loke looking at a clock which reads, say, 7:00 a.m. You can't tell by looking at the clock if it started months ago and has been running ever since, or if someone five minutes ago set it to read 6:55 and started it running. You get the same result either way. The universe is like that clock: It may be 13 billion years old, or it may be that it is a clock created a few thousand years ago to read "13 billion years." (For all we can prove, in that sense, it could be a clock created a second ago, and all our memories before that time were created with it.) Either way, the universal clock reads 13 billion years.
I'll leave out the further impassioned appeals. They won't change anyone's minds; there is now evidence that certain religious beliefs and thought patterns are hardwired into our minds, so what is logical and true for one is not logical to others. Perhaps our greatest need is to be understanding; if you don't believe in evolution, I can only ask that you keep an open mind about what follows, because others are made differently.
For those who want such a thing, I have given an argument (see the note) for why evolution and the Big Bang do not contradict the Biblical account -- but I doubt this helps much. I could repeat that much of what follows is less evolution than simple genetics, which for some reason is non-controversial. But evolution is inherent in genetics: If you have genes, and if you have some genes surviving and some not, you will get evolution. If you want to skip the evolution material, you can derive some benefit from skipping to the appendix on genetics.
Also, evolution isn't just about bacteria developing antibiotic resistance or Homo Erectus turning into Homo Sapiens. It's about anything with a gene analogy being placed under pressure to survive. This has led to a concept known as "universal Darwinism." That is, the belief that natural selection applies to more than just living things. Where there is competitive pressure, meaning that only some members of a population can survive, the larger share of those that survive will be the ones which best meet the selection criteria. It's statistical, and it depends on conditions, and it takes time (quite a lot of time if the selective pressure is slight, which it usually is) -- but it is sure to apply as long as there is some sort of selection. A possible example of this is the meme. Similarly, all else being equal, well-written books will sell better than poorly-written books, so as time passes, libraries tend to contain more and more readable books. Good television shows get renewed; bad ones get cancelled. Politicians who are willing to do and say anything beat those who take stands and cause portions of the population to disagree with them. You could argue that the canon of the Bible, or of other religious books, is an example: If scribes and parchment were infinite, many more books might have been retained in the canon. But since only so many copies could be made, a canon had to be fixed so scribes would not waste their time on relatively unimportant works. And with the canon fixed, those other documents -- the Letter of Barnabas, the Didache, etc. -- effectively died out. The rules of natural selection vary (the primary attribute of a successful television show seems to be stupidity, which obviously is not a very useful trait in nature). But natural selection has to take place.
We might add that evolution as understood today is not really the same as what most of us encountered in school. Part of this, at least in American schools, is the ongoing dumbing-down of the science curriculum for political reasons. But evolutionary theory itself is changing; except for the concept of Natural Selection, what remains is distinctly different from Darwin. The latest great change was brought about by none other than that infamous zealot Richard Dawkins. For many years after Darwin, people thought of evolution in terms of organisms and species. Even after genes were discovered, they were just the mechanism of evolution -- even though genes, once understood, solved a problem that Darwin had never solved: how survival adaptions managed to avoid being diluted by all the un-adapted creatures. (In Darwin's day, heredity was thought to be a sort of an analog process: Children were mixures of their parents' traits. With genes, suddenly, instead of a child getting half of a longer beak, they either had a long-beak gene or a short-beak gene. No half-way measures any more.)
Dawkins further modified this by bringing forward the notion of the "Selfish Gene." The survival imperative applies to genes, not whole organisms. Mostly it is expressed in organisms -- but a gene for blue eyes doesn't benefit directly if it's linked with (say) a gene for light skin; genes can and do evolve and survive separately. It's hard to discuss this without implying purpose on the parts of genes, which is wrong, but it is crucial to understand that each gene is in it for itself; survival of other genes, even of the whole species, is incidental to them. The Selfish Gene view, we note, brings evolutionary biology that much closer to textual criticism. Manuscripts, e.g., do not replicate themselves exactly and in their entirety; they replicate passage by passage and word by word. Usually, if a manuscript is copied, most parts will be replicated. But note the situation is just like a gene: As a gene is either passed on or not, a particular passage is either copied accurately or not. A manuscript may be 99.9% identical to its ancestor -- but any given sentence or word is either 100% identical or it has been changed.
(A side comment on Dawkins. In recent years, he has gone from avowed atheism to open attacks on religion, up to and including his recent publication of The God Delusion -- a book I have not read, and which I think incredibly unfortunate. Having heard Dawkins talk about it, I know that many of his points are valid -- certainly, as one who is trained as a scientist, I know that the American prejudice against anyone who even hints of atheism is strong, ignorant, and unfair! -- but I also know that his book is not going to do any good, and that Dawkins should have known it would not do any good. But we must distinguish Dawkins the scientist -- author of The Selfish Gene and The Extended Phenotype and, at a more popular level, The Ancestor's Tale -- from Dawkins the militant atheist. Dawkins the scientist is a great man. Dawkins the atheist, at best, is someone who completely fails to understand the views of the other side. But we'll ignore that Dawkins.)
A strong note: throughout this article, I use the words "if" and "assume" and "suppose" in the mathematical sense: To say "in the event this is true, that other thing will follow." At no point should this be taken to mean that the "if" statements imply either belief or doubt on a particular point (except the one that is stated above, that evolution is happening now, which is a fact and so need not be questioned).
Also, while what follows will make occasional reference to interesting Biblical parallels, it will pull no punches on evolutionary theory -- e.g. it just assumes humans are descended from apes, which are a branch of primates, which are a particular type of mammals, etc. In fact, there is a stemma of human evolution at one point (mostly because the popular treatments of this are so consistently out-of-date and inaccurate. I can't promise to be accurate, since I'm not a biologist, but my stemma is based on comparing five books by paleontologists, all published within the last ten years as I write).
There are, to my knowledge, at least four scientific/cultural areas which follow true stemmatic models: The origin of biodiversity, the differentiation of human language (and, perhaps, culture), the transmission of written texts, and the transmission of oral texts.
Note that when I say "stemmatic," it means something much more specific than the ordinary use of "evolution" and "evolutionary." The latter can be used for, say, "the evolution of nursing home care" or "the evolution of aircraft manufacture." The latter two are not truly evolutionary; they are merely progressive -- each step goes beyond what is before. Karl Popper and his followers would call this evolution, but it is not in any way the same as what happens in biology. The process of a technology is a series of inventions which do not inherently follow one from the other. A true evolutionary process is one where you start from a small population of types or individuals and watch it increase to a large population of types or individuals, with more diversity, but with each new individual directly descended from some subset of the previous individuals. You also see many if not most of the earlier specimens die out -- and, potentially, you see certain traits die out with them. We must stress that that, in biology, evolution is not progress; it's just change -- or even, in some cases, a lack of change. Evolutionary success, in an ironic way, consists in not evolving; it means a species has successfully found a niche in which it can survive for a long time. Evolution does not have a goal, so it cannot make progress. Whereas aircraft manufacture has a specific goal: Optimizing cargo capacity and speed while minimizing cost of manufacturing, maintenance, crew size, and fuel. The two are simply not comparable.
There are still other processes which are evolutionary -- even stemmatic -- without following the course of biological evolution. Musical genres are an example. Early British folk music has left a number of descendants: Modern British folk in England, New England folk music in, well, New England, southern folk music in the American South. The latter is now called "Country music" -- and it in turn has spawned offspring. The early country is now called "old time country." This was succeeded by "classic country," in which electric instruments began to replace acoustic and the songs were more often composed than traditional. This in turn has been succeeded by pop country, which has very little old-time influence at all, is almost entirely electric, and which is much more centered in contemporary than in timeless themes; almost no one likes both old-time and pop country. The evolution, and even the stemma, is clear -- but the old generations have not died off. Old-time country is not very commercially successful, but it endures, and today's performers are actually more technically proficient than the older musicians. (And, in a deep irony that bears real thinking about, the modern performers, all of whom are deeply devoted to the forms and styles of the past, are consistently liberal, often to the point of radicalism, while the more numerous fans of pop country, a musical form mostly invented in their lifetimes, are the most reactionary conservative population in the United States.) Old music forms are not replaced by new; the market just gets more crowded. This is unlike biological evolution, where all creatures die, and rather unlike even manuscript evolution, where a few last a very long time but most eventually are destroyed.
Even the four processes I specified (living creatures, texts, languages, oral traditions) are different in some regards. Living things, for instance, have genes, which restrict their ability to evolve. A creature with genes can only evolve by Darwinian evolution. Processes without genes can sometimes be altered by Lamarckian evolution (in which they acquire desirable traits due to desire or an external force and then pass them on). This is related to the fact that biological evolution has no (observed) "editor," whereas a text or a traditional folk song can be subjected to deliberate redaction. Then again, where a written text can sometimes be recalled and corrected, an oral text can't. Still another difference is that living creatures can generally reproduce on their own; texts require outside assistance (a copyist). This, in a way, resembles the reproductive cycle of a virus.
Another important point of comparison manuscripts and living creatures is parentage. Creatures which reproduce sexually (which includes almost everything we think of as an animal, and most large plants) have two parents. They have inherently mixed ancestry. This contrasts to most of the other processes. Languages are rarely very mixed (though English, with its Germanic base and its heavy Norman French overlay, is in fact a mixed language). Biblical manuscripts are, almost without exception, mixed, but the mixture is intermittent: Most of the transmission stages in a manuscript's ancestry invole only one parent, but at various times, individual generations will have two, as a text is corrected from a different manuscript than the one used to copy it. This is unlike animals, but quite like a lot of plants, such as strawberries or elms: They sent out clones of themselves, by runners or the like, but also reproduce sexually at times. (It has been speculated, though I don't believe it's been proved, that most of these dual-reproduction species use the two methods differently: Cloning is for populating the local area as much as possible, since the plant's own genes have been successful in that environment. Sexual reproduction, which mixes genes, is for sending seeds out far and wide, where a different combination of genes might be more successful.)
Thus every one of the four processes has its own characteristics, and the differences are often much more important than the similarities. Textual Criticism, for instance, is trying to look back to the original document. Evolutionary biology seeks to explain present creatures and perhaps predict what will come next. But each can, potentially, inform the others. There are plenty of parallels, from the crucial to the trivial (e.g., just as old manuscripts are sometimes cut up to be sold in pieces to collectors, sometimes fossils are broken so that the fragments can be sold in pieces to archaeologists).
I will admit that my thinking about this was sparked by reading The Ancestor's Tale by Richard Dawkins. I had already known that biological evolution had parallels to textual -- but Dawkins kicked up so many interesting analogies that I decided it was time to bring them into TC as best I could. Which probably isn't very well, since my training is in physics, not biology. But maybe it will point the way for someone else.
Where possible, I express as what follows as questions, since in some places the parallels are partial, and in others no one has even looked for parallels.
I already noted one of the key points about biological evolution: That it proceeds through genes. Genes are, in some regards, crucial: A person whose parents both have type O blood, for instance, cannot have type A or type B blood; the genes aren't there. And the only way one can create a new blood type -- "Type C," let us say -- is by mutation. Such dramatic mutations are much more likely to be fatal than to give rise to something new and useful, or even new and neutral.
Are mutations, then, the analogy to scribal errors? A lack of analogy lies in the way they are treated. A single scribal error will almost certainly not be fatal to the manuscript. The possibilities are not that it will be kept or thrown out, but that it will be corrected or it will not. If it is corrected, the error will not propagate. If it is not corrected, it still may not propagate -- if it makes nonsense, it will be replaced with something else, though this correction may not be the same as the pre-error reading.
Genetic mutations have another interesting trait: Those which survive come in two types. Some mutations are "founder mutations," others are "hot spot mutations." A "hot spot mutation" is a gene which is particularly susceptible to damage, in which case the mutation can happen again and again. The disease achondroplasia, which results in dwarfism, seems to stem from this. Hemophilia, too, apparently -- the hemophilia gene Queen Victoria spread around Europe is believed to be a mutation she herself suffered, since none of her parents' relatives appear to have been carriers.
Other mutations are believed to have happened only once, with a "founder"
who passed them on. Sickle cell trait is a likely example of this: Someone developed
a single copy of the gene -- and was advantaged, because sickle cell trait helps
prevent malaria. So that person (probably African) left many descendents (none of
his or her children would have sickle cell anemia, note, because the other parents
were free of the trait. The first chance for sickle cell disease, as opposed to
sickle cell trait, to appear would have been in the
grandchildren, and that only if siblings mated; in practice, it was probably at
least four or five generations before the first child with sickle cell anemia
appeared. And by then the gene was widespread -- and odds are that no one could
tell where the disease came from, even assuming the culture believed in inheritable
disease, which few did).
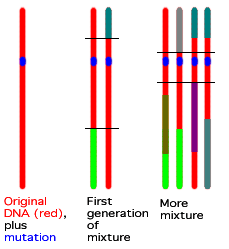
Biologists now are using founder mutations to try to trace the age of particular traits. They do this by comparing the DNA around the mutated site: the more similar DNA the carriers of the trait have, the more recently it must have developed. The illustration at right shows this: The first strand of DNA (the mostly-red vertical line on the left) is the DNA of the original chromosome in which the mutation happened. Red DNA (plus the blue mutation) comes from that original mutated individual. DNA of any other colour is from someone else. In generations following the first, the chromosomes mix, with more and more non-red DNA becoming part of the daughter chromosomes. But they mix only slowly, with typically only a few mixes per chromosome per generation; most genes stay with their chromosomal neighbors in any given generation. As time passes, you have more and more swaps and less and less of the DNA near the mutation stays with it. (The black lines in the drawing show the common DNA in each generation. Note how much more there is in the middle generation than the final generation.) By comparing the DNA of several people with the mutation, and seeing how much DNA they have in common, one can estimate the age of the mutation. Textual critics do not have this sort of "clock" for variants, but certainly the types of variants will be familiar. "Hot spot" variants are those which could occur to scribes individually -- h.t. errors would be an obvious example. Assimilation of parallels could also occur this way (this may even be the reason for the claimed existence of the "Cæsarean" text-type), and maybe the expansion of Christological titles. But a radical change like adding the longer ending of Mark must be a "Founder mutation" -- someone wrote it, and it stuck. (Note incidentally that we cannot always tell which type a variant is just by looking at the variant: In the case of Mark 16:9-20, it is a founder mutation if the longer ending is not original, but if it is original, and was deleted, it might be a hot spot mutation, lost in several copies of the book since it was near the end of the scroll. In biology, we can usually tell which form is original. Perhaps less so in textual criticism.)
(Even ancient founder mutations, which are old enough to have very little context left, can sometimes be dated -- in mitochondrial DNA. Nicolas Wade, in Before the Dawn, pp. 106-107, tells of the work of Martin Richards on populations in Europe. He created a family of DNA mutations -- a relatively straightforward task -- and then noted the order in which mutations "joined." This gives him the ability to approximately date them. This technique could presumably be used in dealing with unmixed local texts. It is less obvious how it can be used in mixed texts. The fact that Richards applied his technique only to mitochondrial DNA, which does not mix, is significant.)
The obvious analogy to the splicing together of chromosomes is block mixture, though there might also be some analogy to the case of different scribes writing different parts of a manuscript. They will at least make different sorts of spelling errors, even if it doesn't affect the text-type.
There is another key similarity between genetic and textual mutations: A change in a single gene in the genome of a living creature doesn't affect anything else (at least directly). Similarly, a change in a particular reading does not change the rest of the overall text. (This is unlike language, where a change in grammar at least will affect the whole language, and also unlike oral tradition, where the loss of a line or two of text may force a major reshaping.) The real difference lies in the fact that genes have boundaries; texts don't. Scribes copy in all sorts of ways -- letter by letter, word by word, phrase by phrase. Something that is a genetic unit to one scribe won't be to another.
Still, the fact that scribes copy in pieces brings us to an interesting point. Different stretches of text can in fact have different ancestry. The family tree of a passage need not be the same as the family tree of the manuscript as a whole.
Dawkins refers to this as the "historical" and "genetic" ancestors.
Turning to one of our parallel disciplines, we can turn to the folk ballad known as "Barbara Allen." This is probably the best-known song in the history of the English language, having been collected roughly 1000 times (and as a result, I've used it in several examples, notably in the article on oral transmission). Two folklorists, Charles Seeger and Ed Cray, once set out to examine its ancestry. Seeger looked at the tunes, Cray the texts, and they did not compare their results until they had done their basic classsification.
Seeger found that the song had four basic tunes. Cray found that it had four basic text-types. It is logical to assume that the two arose separately -- that is, at one time, text-type A had a particular tune associated with it, and text-type B had its own tune, and so forth.
Not any more. Texts and tunes are completely dissociated. (It has been shown in other contexts that people frequently swapped tunes for songs.) Although any particular version will presumably go back, without much mixture, to one of the early song groups (the historical ancestor) it may well have derived its tune from another group (the genetic ancestor).
To take an example of how this works in biology: Most genes are passed via mixture: You can get them from either parent, and so you can't tell, just by looking at the end product, which ancestor supplied a particular gene. But there are two exceptions: Mitochondrial genes, and Y chromosomes.
We already mentioned mitochondria once; let's talk about that a little more. Mitochondria are the cells-within-cells that supply the power used to run the rest of the cell. They are found in all cells -- but the father's sperm don't transmit them to children; only the mother's eggs do that. And mitochondria have their own DNA -- just a handful of genes, but they are genes. So every child gets his mitochondrial DNA from his or her mother, and from her mother before that, and her mother before that, and so on forever. Theoretically, it all goes back to a hypothetical first mother. (In fact, it appears that there is a first mother, from whom all human mitochondria are descended. She is called "Eve," naturally, and is estimated that she lived about 140,000 years ago.)
Incidentally, the tracing of mitochondrial DNA gives evidence, stemmatically, that the origin of humanity was in Africa, or at least that there was a choke point in Africa from which all later humans descended. This is shown by a stemma first published in Nature in 1987 (I found it printed in James Shreeve, The Neandertal Enigma, p. 56). It's so complicated that it is printed as an arch, with 135 data points arranged in a tree. But let's take only its first few branches and see where they lead. Note that each branch is a genetic variant: Either your mitochondrial DNA has one reading or it has the other. Barring coincidental agreement (i.e. two pieces of DNA suffering the same mutation, which is possible but unlikely), this gives an absolute stemma. At the end of each branch, I show the continents where it is found: Af (Africa), As (Asia), Au (Australia), NG (New Guinea), Eu (Europe).
"Eve"
|
--------------
| |
A B
| |
| -----------------
| | |
| C D
| | |
| | ----------------------------
| | | | |
| | E F G
| | | | |
Af As Au Af,As,Au,Eu,NG Af,As,Au,Eu,NG
Now it must be stated that this stemma admits of two possible interpretations, depending on how we root the stemma: First, that the B version is the original, probably in Asia (since that's the place common to C and D), with humanity spreading from there; second, that the original sequence (which could have been A or B; we cannot tell) originated in Africa, and split there, with only a small population of B migrating into Asia and hence into Europe, Australia, and so forth. This is, ultimately, the Bédier Problem of a two-branch stemma -- but in this case there is so much variation in the descendants of A that the biologists think the point of origin must have been in Africa.
Just as mitochondrial transmission is exclusively in the female line, the Y chromosome is exclusive to males, meaning that any man got it from his father, and his father's father before that, and so on back to the beginning. As with mitochondrial DNA, the divergences in its DNA can be diagrammed. This points to an "Adam" who lived about 60,000 years ago.
To console the creationists, we note incidentally that this does not automatically mean that there cannot have been a contemporary Adam and Eve, though it obviously seems unlikely. The Biblical Adam and Eve, if they existed, were the earliest common ancestors of all humanity. The "Adam" and "Eve" of the molecular biologists are, respectively, the last common male ancestor of all males and the last common female ancestor of all humans. There is no inherent reason they should live at the same time. The following genealogy shows how this could happen, where s01, s02... represent unnamed sons, d01, d02... represent unnamed daughters, and === represents a marriage.
Eve === Adam
|
----------------------------
| | | |
Seth === d01 s01 === d02
| |
---)--------------------------
| | | |
----------)------------)-------- |
| | | | | |
Enosh === d03 s02 === d04 s03 === d05
| | |
...all produce many more children...
Now observe: If you trace all the children back in female line, some are descended from d01 and some from d02. Thus their most recent female ancestor is Eve. But every male offspring is descended from Seth, via Enosh, s02, or s03. Their most recent common ancestor is Seth, who lived later than Eve. This does not mean that s01 had no descendants; he has as many as Seth. But, because they were all daughters, his y chromosome is extinct; only Seth's is preserved. In fact, were this genealogy correct, Seth's X chromosome would also be extinct, because he had only sons, but we can't prove that because we can't trace X chromosomes as we do Y.
For that matter, we would note that, if the Biblical account is literally true, then the last common male ancestor cannot be earlier than Noah (since all males who survived the flood were his sons), but the females must go back some time before that. Personally, I find that kind of spooky.
This has other implications for Biblical history: Dean Hamer's book The God Gene, chapter ten, "The DNA of the Jews," starting on page 180, tells of a study of the Jewish "Cohens" -- the people who claim to be the descendents of the ancient Aaronite priestly family. The study seems to reveal that they really are a family: based on the Y chromosome data, the "Cohens" almost all derive from a common male ancestor who lived at least two thousand and possibly as much as 3250 years ago. In other words, the modern Jewish priestly lines go back at least to the end of the Second Temple era, and possibly even to the period of the Judges or the end of the Exodus. We can't prove that the common ancestor was Aaron or Eleazar -- for all the data proves, it could be Simon Maccabee or even, theoretically, Annas or Caiaphas -- but it proves that these people are indeed descended from one common ancestor.
We can go beyond this. Just as, ultimately, there can only be one autograph of a New Testament book (or, at least, only one first draft, since the author may have published multiple editions), there must -- by virtue of the fact that humans have not always existed -- be a human being who is an ancestor (not the ancestor, but an ancestor) of all future human beings. This process has been studied a great deal; Dawkins talks about it on pp. 42-44 of The Ancestor's Tale. It can also be shown that, if we go back a certain number of generations, every person (and so every manuscript) is either the ancestor of all manuscripts or is an ancestor of none -- and, in a typical population, more are ancestors of all than are ancestors of none. We'll see more implications of this below.
Most reproductive processes have this phenomenon of earliest and latest common ancestors. For mammalian life, it's likely that the earliest recognizable ancestor is some ancient creature that was the first eukaryote (that is, the first being with a clearly delineated nucleus, mitochondria, and other characteristics of modern cells). But the common ancestor even of such seemingly-unrelated creatures as human and trees is probably more recent than that -- in the case of, say, humans and apes, it's much more recent.
Similarly with languages. German and English, for instance, have a known common ancestor in proto-Indo-European, thousands of years ago. But their most recent common ancestor is Old Germannic (or whatever you want to call it), and the two diverged only about 1500 years ago. And even after the divergence, they were still mutually comprehensible for some hundreds of years, and probably swapped a few new words.
In the case of literary texts, the analogy is in this case quite exact: the earliest common ancestory is the autograph, while the latest is the archetype. The analogy is precise in another way, too: You can only reconstruct back to the latest common ancestor. The ancestors of the Indo-Europeans surely spoke some language. But we can't reconstruct it (at least until we can bring still more languages, which diverged earlier, into the Indo-European family tree). In the case of language, we can't even prove that there was a single original language; many linguists think there was, but this strikes me as a little strong; the divergence in styles of language is so great that it strikes me as quite possible they came about after the earliest population had split (i.e. the language capability was there but initially unused). With such uncertainty, we surely cannot recreate the "original language." All we can do is recreate proto-Indo-European, the language as it was spoken at the time of the first split in the family. Similarly, if the archetype is more recent than the autograph of a particular literary work, you can only reconstruct the archetype; beyond that, all is conjecture.
We note incidentally that the reconstruction of language can often interact with other rather stemmatic disciplines. Staring in the early 1960s, Joseph H. Greenburg began a systematic attempt to link all languages together into families. His success has been mixed; very many linguists now accept his results for the "Afroasiatic" superfamily (consisting of the Egyptian, Omotic, Cushitic, Berber, Chadic, and -- notably for our purposes -- Semitic families), but they are less sure of his other results. The interesting point is that his results have often been correlated with other sorts of data, such as DNA evidence of people. (See Nicholas Wade, Before the Dawn, pp. 218-232.) I find myself wondering if we might not learn something by trying, say, DNA analysis of the parchment from various manuscripts.
Population genetics has had other interesting results which might affect our analysis of the New Testament tradition. A detailed comparison of the population of Iceland showed two interesting things, both of which affect the claim of "normal transmission" proposed by the advocates of the Byzantine Text. One is that not all populations reproduce at the same "generational speed" -- in Iceland, the average interval between generations was 29 years for females, 32 for males. Presumably this is because females marry younger, but still, populations clearly do reproduce at different rates. In addition, the work of DeCode Genetics has shown that 92% of Icelandic women born since 1972 were descended from just 22% of the women born 1848-1892, and 86% of men born since 1972 were descended from just 26% of the pre-1892 group. It has long been suspected, mathematically, that populations reproduced this way; now it has been shown empirically (see Wade, p. 244).
But it needs to be repeated that "common ancestor" is a sort of a relative term, because genetic and historical ancestors may be different. This is true in two different senses. One is genetic reduction: A human being or other creature inherits only half his chromosomes from either parent. So if our Ancestor is A, his child A1 has exactly half his chromosomes. A1's child A2 has only, on average, half of half, or a quarter of A's genes. A3, the child of A2, has only an eighth, A4 has only 1/16, A5 has only 1/32, A6 has only 1/64, A7 has only 1/128, and so on.
But note that human beings have only 46 chromosomes. 1/64 of 46 chromosomes is less than one; 1/128 of 46 chromosomes is less than half a chromosome. In other words (barring reinforcement by inbreeding), after seven generations, odds are that an offspring shares no chromosomes with any random ancestor. The same might well be true of a text that has been through enough stages of mixture: It might have an Alexandrian ancestor, say, but after enough Byzantine corrections, it's no longer possible to tell. Each generation of mixture has taken more of its Alexandrian-ness away.
(Note: The above statement is very oversimplified. Chromosomes in fact are not conserved across generations. As noted above, except for sex chromosomes in males, the chromosomes swap genetic material from generation to generation. This is one reason why human chromosomes are so chaotic, with parts of singles genes scattered all over the place. But while this is very important for actual evolution and molecular genetics, it concerns us not at all. The only thing we need to remember is that one can be descended from a person without having a single gene derived from that person. It sounds insane, but it is not. The relevance is this: Manuscripts which are the result of mixture reproduce in the same way as genes: Some parts come from one parent and some from another. Given enough generations, there may be no readings left which derive uniquely from the ancestral manuscript. An Alexandrian ancestor can be mixed to the point where its offspring becomes purely Byzantine, or vice versa. It doesn't even take that many phases. Given the level of correction found in, say, C, it would take only about four generations to go from Alexandrian to Byzantine.)
The other part about genetic and historical ancestors can perhaps be illustrated by using blood types again. Human beings have two major types of blood clotting factors, A and B; the absence of these gives type O blood.
But, interestingly, it has been shown that chimpanzees also have type A-B-O blood. I've also seen it stated that gorillas have A-B-O blood, but I haven't seen this confirmed. I've found the A-B-O split in chimps stated in several places. (Matt Ridley, on page 26 of The Agile Gene, states that chimps have types A and O blood, while gorillas have type B. The stemmatic implication, obviously, is that the common ancestor of humans, chimps, and gorillas already had types A, B, and O.) No matter what the exact distribution of clotting factors, it is clear that the evolution of the factors predates the split between humans and chimps, thought to have taken place about six million years ago. If the report about gorillas is true, then the A-B-O split took place one or two million years before that.
Now compare this to the dates for "Adam" and "Eve" given above. The blood type split, since it goes back six million years, is at least thirty times older than Eve. And, as regards blood type, one can be more related to a chimp with one's own blood type than with a human of a different blood type. For example, I am type O. I share the genes for clotting factors with all type O chimps -- and I don't share them with you if you are type A, B, or AB. (Note: Because type O is a recessive, that's not entirely true. If you are A or B, I may share one type O gene with you. But I may not. And if you're AB, then I know I don't share any genes with you.) Although I am historically descended from human beings and not from chimps, as regards blood type, I am more closely related to some chimps than I am to some people.
Note that, again, this does not require some descendant of "Eve" to have mated with a chimp or the like. If "Eve" had, say, A-O genes, and "Adam" had B-O, or Eve and Adam had OO and AB, or vice versa, then the blood types could all be descended directly from them. That's the whole point: We aren't talking ancestry but common genes. Because of the genetic reduction -- which can become genetic drift -- with regard to a particular gene, I may be unrelated to "Eve" or "Adam" -- or even both.
An even more obvious example of this is the distinction between male and female. Different creatures have different ways of determining sex -- among some fish and reptiles, it's dependent on environment, e.g. But in mammals, it's genetic. There are a handful of Y chromosome genes which cause maleness by turning on a few switches. This is universal in all mammals. Thus, if you are male, the basic genes to make you male have to go back all the way to the earliest mammals, if not earlier. In regard to those genes, you are closer to a male rabbit or horse than you are to a female human. Patently absurd in an overall way -- more than 97% of a male's genes are found in female humans, and they aren't found in horses or rabbigs -- but true for those specific genes.
The analogy to text-types is interesting. If a manuscript mixed in a foreign element, that element is more related to the source of the mixture than the historical ancestor. So, for example, "John 7:53ffff." probably originated in the "Western" text-type. But it's found in some manuscripts which are otherwise mostly Alexandrian. Chances are that it was introduced by mixture, and that that passage, even in L or 579 or whatever, is genetically "Western," not Alexandrian.
This invites a very different way of looking at stemmatics. We tend to think of genealogy in a top-down sense, with an autograph, then various copies, and copies of the copies, down to the manuscript. And we may think of the manuscript as being derived from a variety of sources going back to the original -- a bottom-up approach.
But in light of DNA descent, we might have to think of this in different terms. Each manuscript has a historical descent which takes it back to the autograph. But each reading can have a different descent.
This may sound a bit like the Alands' local-genealogical method. It is not. A better analogy might be to a train: Each car on the train comes from a specific place -- but not always the same place. Indeed, each of the cars may contain packages from multiple sources. The model we see is much more three-dimensional than our traditional view. Each reading has a specific path back to its origin (which may be the autograph or may be a corruption). The diagram below may give some idea of this. There are three text-types, called by Greek letters but shown as red, green, and blue. At the bottom we have a particular manuscript with a particular set of readings, all derived from one or more of the three types.
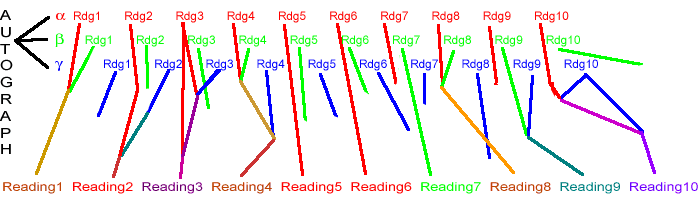
This picture can make life look despairingly complex. Complex it is, but there is hope. Because we can demonstrate that every manuscript has a common ancestor. Somewhere. It may be the autograph. But it's there. Similarly, every reading has a common ancestor.
Take each readings in the final manuscript individually. Reading 1 is has red and
green ancestry. Reading 2
is red and blue. So is reading 3. Reading 4 has influence from red, green, and blue.
Reading 5 is pure green, reading 6 pure red. If we chart this, we find that each of the
text-types contributed to certain readings:
RED: Readings 1, 2, 3, 4, 5, 6, 8, 10 (total of 8 readings)
GREEN: Readings 1, 4, 7, 8, 9 (total of 5 readings)
BLUE: Readings 2, 3, 4, 9, 10 (total of 5 readings)
Note that no type contributed to all of the readings. But consider red, with influence in eight out of ten readings (including two, #5 and #6, which are purely red). It is safe to call this a "red" manuscript. When we are in doubt about a particular reading's origin, it is not a bad bet to assume that it's red, though the matter is not certain. We just have to understand that a manuscript of a particular type may have readings not of that particular type -- as a person of Scandinavian origin might have inherited a few genes which go back to the Middle East or China.
This is the solution of the seeming paradox above, that, if we go back a certain number of generations, every manuscript is either an ancestor of all manuscripts now surviving, or of none, and more were ancestors of all than of none. Of the first few dozens copies, most -- if not destroyed without being copied -- would have influenced surviving manuscripts. They would be historical ancestors. But would they have contributed significantly? Probably not. It's quite possible that a manuscript of a separate text-type (say the Greek that underlies one or another Latin or Syriac version) could have influenced all our modern manuscripts -- and not have a single one of its unique readings survive! Obviously this could affect how we view our particular collection of mixed manuscripts....
To this peculiar-sounding proposition we can also compare the ancestry of books in a corpus. The earliest copies of the Pauline corpus, for instance, came together before all the books were accepted as being part of it. (Hebrews obviously was not included in some early copies; compare also the contents of P46.) Might books which circulated on their own and were eventually added to a corpus have gone through a history unlike the other books in the later copies of the corpus?
This brings us to another problem that doesn't get much thought. Most people agree that text-types exist, though they disagree on how to use them. But when and why did they come to exist? This is more complicated than saying, "The Byzantine text was in existence by the fifth century." If we ignore the effects of mixture, there was some point at which any two text-types split -- that is, there was some manuscript of which we can say, "This is the ancestor of all Byzantine (or Alexandrian, or Western) texts, and it is not the ancestor of all non-Byzantine (or non-Alexandrian, or non-Western) texts." But that manuscript will be closely related to other texts of which this is not true. It may be the ancestor of all Byzantine texts, but it is not itself characteristically Byzantine! So should it be called Byzantine?
Dawkins gives an analogy of this in terms of the transition from monkeys to apes. The usual distinction between monkeys and apes is that monkeys have tails. At some point, a female monkey bore an offspring which would be ancestral to all modern apes (and humans). But that ancestral creature, the offspring of a monkey, surely had a tail. It was, to all appearances at least, a monkey -- and surely still capable of breeding with other monkeys. At what point did that lineage become apes? When the tail was lost? What if the tail was lost gradually -- there are a lot of monkeys with short tails. There is no answer to this. It merely teaches us an important rule about evolution: That trends and processes are more inportant than the staging points along the way. We call the ancestors of humans by names such as "homo habilis" and "homo erectus." But these actually represent more a trend than anything else -- a trend toward bigger brains. There is no place or time at which you can halt and say, "This is completely different from everything that came before or after." Does this affect how we use manuscripts? Probably -- because every manuscript, except the autograph, is just a stage in such a process. It's a fossil representing a stage in the history of transmission. It is not the beginning, and not the end. The goal is presumably to see where it points.
Another interesting analogy to Paul exists in the case of the human chromosome 2. It is a curiosity that apes consistently have 48 chromosomes, but humans have only 46. It is now pretty well established that the reason is that our chromosome 2 (which is, of course, the second largest in the genome) is in fact a combination of two shorter ape chromosomes. In this sense at least, humans are different from apes. Compare this to 2 Corinthians, which certainly appears to be fragments of two or three or four letters squished together.
The assembly of books into a corpus, or of separate letters into a sort of a whole, incidentally has another evolutionary parallel, though this is much more speculative. One of the great problems of biology is to explain how the current system of our cells came to be. We have a controlling mechanism, the DNA, which stores the information needed to create and control a cell -- but which is useless for anything else except information storage. We have a bunch of proteins, many of them enzymes, which can actually do things, but which cannot store information or reproduce. For life to come about, at first glance, it would appear that these two would have to come into existence together -- the so-called "irreducible complexity" argument.
Which is, however, false. The argument is false in general, because it ignores the usual trend of evolution, which is toward simplicity (that is, natural selection sheds all unneeded features; if something is truly irreducibly complex, odds are that it is at the end of a long chain of evolution based on less effective, more complex methods). And it is false in this particular case because it appears to ignore how life started. The goal is to find a "naked replicator" -- the original molecule that could make copies of itself. Neither DNA nor proteins fit this description; they're too specialized.
But there is another molecule found in nucleii: RNA. While we can't prove it (yet), it appears likely that RNA was the original replicator. Unlike proteins, it can reproduce -- although inefficiently and with a lot of errors. Unlike DNA, it actually has some ability to act on its environment -- indeed, the ribosome, which converts DNA sequences to proteins, contains RNA as well as protein, but no DNA. And it is RNA, not DNA, which transmits the actual sequence of proteins to be encoded.
This is notably significant, because most of this process has been recreated in the laboratory -- that is, people have recreated the conditions on the early earth, and added energy. The first experiments of this type, in the mid-twentieth century, quickly yielded amino acids. More recently, we've started to move toward an actual RNA factory (see Matt Ridley, Genome, pp. 18-21; in addition, Dawkins, in The Blind Watchmaker, p. 190, tells how Manfred Eigen evolved RNA in the lab without any influence from existing life, though he started with raw materials slightly more complicated than just the carbon dioxide, water, methane, and such of the amino acid experiments. According to John Maynard Smith, The Theory of Evolution, pp. 9-11 of the 1993 Canto edition, an even more interesting aspect of this process is that, when run several times, it produces similar but not identical end results, showing that natural selection does operate to favor the same types of results but not the same exact results). Add it up and it's clear that humanity has recreated something pretty close to a modern virus: Some viruses (called, for obvious reasons, RNA viruses) have no DNA, just RNA and protein. These viruses probably don't go back to the days of the "RNA World" (there wouldn't have been anything for them to infect), but they are very likely similar to life forms of that era.
Incidentally, Dawkins, The Ancestor's Tale pp. 576-578, describes an experiment undertaken by Sol Spiegelman on the RNA of a bacteriophage (virus which infects bacteria) called Qb. Spiegelman not only caused its RNA to reproduce without a bacterium (indeed, without parts of the original virus), but he induced it to evolve. The final RNA strand was only a seventh of the size of the original, and no longer had the genes to invade bacteria -- but it could reproduce faster and more effectively in the lab setting Spiegelman had created. It was, by any rational definition, a new species.
Turning back to ancient history -- having created those RNA replicators, the next step must have been for them to group themselves together. After all, one RNA replicator can only perform one or two functions apart from replicating. But if they group, they can combine to perform multiple functions. This is the analogy to the collecting of books in a library.
And once they are grouped, then it becomes possible to branch out. RNA is not a very good replicator. It's unstable, and inefficient at duplicating, and while it can influence the outside world, it can't do so very easily. A more stable replicator and a more complicated set of enzymes would work better. With a bunch of RNA working together, so that some could do the work of surviving while others experimented with replicating, it was possible to make the relatively minor chemical changes needed to turn RNA into DNA. And once that was done, the replicator system could truly take off.
Allow me an analogy. Think of life inside a nucleus as being like a computer. There is memory (DNA) and input and output devices (proteins). It might seem that a computer could not exist without both -- that both had to come into being at together and all at once.
But it's not true. Think of the first American computer, ENIAC. It didn't have a display as we'd now think of it, nor a keyboard or mouse, and it didn't have the huge collection memory chips a modern computer possesses. A task was actually wired in, and computed, and the result read off of the state of the vacuum tubes, or something equivalent. This is the computer equivalent of the RNA universe: It doesn't work very well, but it works well enough that you can start fiddling with it, adding on a serial output rather than a bunch of blinking lights, adding a few memory cells to allow stored calculations, and so on. Once you have something that works, it can be improved.
The key in all this is the grouping: There was one sort of evolution before RNA molecules came together. It was all very different afterward.
Incidentally, since we're talked about human and ape chromosomes, we can show how stemmatics affects evolutionary arguments. Recall that humans have 46 chromosomes, while apes have 48. How do we decide how many chromosomes the last common ancestor had? If humans and apes split separately (that is, if we have a binary tree, with apes on one side and humans on the other), then we would have no way of knowing. But it appears on other grounds that the correct stemma is as follows, with surviving species shown in bold and their number of chromosomes in parentheses. (Note: There is not universal agreement on this. Richard Leakey and others still seem to hold to the view that homo habilis is not derived from the australopithecines but is their contemporary. But he is in the minority, and his theory requires a much more complicated genealogy with more "missing links." I don't think it will hold up. There are lots of other minority theories, too -- one would dissolve homo habilis. Another would reclassify homo habilis as an australopithecine, which shows mostly how hard it is to define different genus. Many would now subdivide homo erectus into homo ergaster and homo erectus, with the former being perhaps the ancestor of modern humans and the latter an extinct offshoot. Many would put homo heidelbergensis between erectus/ergaster and the sapiens/neandertal split. Consider the stemma below simplified: These are the parts almost everyone agrees on.)
ancestral ape (sivapithecus?)
|
--------------------a-----------------------------------------------------
| |
| |
b (dryopithecus?)------------------------------------------ |
| | |
| | |
| | |
g---------------------------------------- | |
| | | |
australopithecines ("Lucy") proto-chimp | |
| | | |
---------------------- ----------d------- | |
| | | | | |
gracile robust | | | |
australopithecines australopith. | | | |
| | | | |
homo habilis | | | |
| | | | |
homo erectus | | | |
| | | | |
------------- | | | |
| | | | | |
homo neandertals chimpanzee (48) bonobo (48) gorilla (48) orangutan (48)
sapiens (46)
Now look at the five surviving species: us (homo sapiens), chimpanzee, bonobo, gorilla, orangutan. I've labelled four ancestors: a is the common ancestor of all surviving apes and humans. b is the ancestor of humans, chimps, and gorillas. g is the common ancestor of humans and chimps. And d is the common ancestor of chimps and bonobos. It is evident that d had 48 chromosomes. g is not certain if we look at the offspring -- but we note that d and the gorilla both have 48 chromosomes. So b must have had 48 chromosomes, and it is highly likely that g had 48 chromosomes. And so did a. Hence, 48 chromosomes is the norm for apes. Somewhere between g and ourselves (that is, in the last six million years or so), two ape choromosomes combined to form our chromosome 2. (I've heard it speculated that this was the moment at which humans became human, that is, the creature made in the image of God. Hard to prove that scientifically....) We don't know when it happened -- and our only DNA samples come from the post-homo erectus stage, which is probably at least a million years too late to be useful -- but it happened somewhere. It might be logical to assume that it came at the homo habilis stage, since that's considered a new genus, but again, nobody really believes those genus designations, so that's no help. Rather than speculate too much, I'm just trying to note how stemmatics and evolutionary biology use exactly the same rules.
Let's go back and consider one more point regarding genetics and blood types, as regards Adam and Eve
and the dominance of the Byzantine text. If one assumes that all blood types
go back to the ancestors, then the Biblical Adam and Eve (if they existed)
must, between them, have had the genes for A, B, and O types. We noted a couple
of ways this could have happened above. There are only a handful of possible genetic
possibilities to allow this situation to survive. The list below shows the two
genes possessed by each of the two parents. (It doesn't matter, in this case,
which is Adam and which Eve; just that one parent has each of the gene combinations
shown below.)
AA BO
AB AO
AB BO
AB OO
AO AB
AO BO
Each of these sets of genes would, if perpetuated equally, produce certain
ratio of A to B to O blood types in the children.
Taking the last, for instance,
we have 25% A genes, 25% B genes, and 50% O genes.
Recall that the O gene is recessive;
the only way to have type O blood is to have OO. A and B are dominant, so AA and AO
both give the offspring type A blood; similarly, BB and BO both yield type B blood.
So ancestors with the combination AO and BO would yield the following
expected values for the prevalence of types A, B, AB, and O blood:
A: 5/16
B: 5/16
AB: 1/8
O: 1/4
This, note, is the highest possible probability for type O (or any other
recessive gene) when there are only four ancestral genes available: One in four,
or 25%. Any other combination of genes in the ancestors will give only one person
in 16 with type O blood. No matter what the genetic situation, type O should be rarer
than A or B. In fact, we see no such thing: Type O blood is the most common blood type.
Different populations vary, but probably about half the world's people have type O
blood, roughly a third have type A, an eighth or so have type B, and a few odd
percent have type AB. We don't know how this situation arose, but presumably it's
in response to some sort of circumstances which have caused differential
selection of blood types.
There is also the curiosity that some populations have very different type ratios than the overall world population -- e.g. Scandinavians, I've heard, have a much higher than average rate of Type A blood than the average. Type B blood is never common, but in parts of central Asia, 30% of the population is Type B. On the other hand, Type B blood is so rare in American Indians that it seems likely that it it all due to recent contamination by Europeans. Another blood factor, the Rh- type, is found in only about a tenth of the world's overall population -- but hardly ever in native Africans, Asians, or Australians. All Rh- blood seems to be derived from Europe, with the most extreme population being the Basques, a third of whom are Rh-. (This to me raises the interesting question of whether Rh- blood might somehow be related to the Neandertals, who were entirely European and lasted longest in areas fairly near where the Basques now live, but that's entirely another issue.)
Remember that A-B-O, at least, predate the rise of humanity. (I'm not sure about Rh factors.) That means that the ancestors of any population will have had access to A, B, and O types. Yet there are no Indians with type B blood, as noted, and the various tribes have different ratios of A to O type blood, with extremes of more than 80% of one type or the other. This is an example of the well-known phenomenon of "genetic drift" -- in a small population, a particular gene can simply drop out of the gene pool. (Nicolas Wade, Before the Dawn, p. 117, reports on a study which appears to show that certain tribes were genetically isolated for seven thousand years!) Clearly when Native Americans came to North America, they were so small a population that Type B blood died out, and in the closed circles of some of the other tribes, types A and O came close to doing the same; it may well be that the handful of people with A or O blood in those tribes are due to incidental recent contamination. (This, incidentally, may be one reason why Indians were so easily stricken by European diseases: They had fewer "polymorphisms" of blood and other traits. According to Matt Ridley's book Genome, p. 141, type O blood confers some slight resistance to malaria, which could explain why it is relatively rare in Scandinavia: malaria has never been an issue there. A and B, on the other hand, while they leave people more vulnerable to malaria, improve resistance to cholera, with the handful of people with AB blood being nearly immune. Havingall three blood types around thus confers immunity to a wide range of diseases -- not directly for individuals, but for the population as a whole, and if enough people are immune, it's harder for a disease to take hold. A polymorphism is simply a case of different characteristics among different individuals, making it harder for a particular invader to prey upon all of them. This particular aspect of genes and evolution probably is not of textual significance, though it's perhaps an argument for not following one text-type slavishly, because the textual equivalent of genetic drift will almost certainly cause at least some good readings to die out in all text-types.)
The moral is clear: Both overall distribution of blood types, and the distribution within local populations, show different types becoming more or less common. One simply cannot assume, as Byzantine prioritists sometimes do, that texts reproduce "normally." It may happen sometimes -- but it must be demonstrated, not posited.
Looking at genetic differences between species brings up other interesting points. It should be noted that there are a lot of genes that function in almost exactly the same way in all mammals, from mouse to human. A mouse muscle cell does much the same thing as a human muscle cell; the difference is not how muscle cells are made but how many of them each creature has and where they are located. Similarly with very many other genes. (This probably explains why mice and humans have about the same number of genes. Most genes are muscle genes or blood genes or brain genes or liver genes, for making muscle or blood or brain or liver cells; humans and mice make the same parts. There are only a few genes -- the Hox genes and a few like them -- that tell the body where to put a muscle cell or a brain cell or a liver cell. And it doesn't take more Hox genes to say "make a huge human brain" than to say "make a small mouse brain"; it just takes different ones, calling for different numbers of cells. Hox genes are found in just about every animal tested, from humans to mice to fruit flies, and they work in almost the same way. And even a fruit fly, which has something like a two millionth of the mass of a human being, has about a quarter of the hox genes that a human has.)
We might well also compare the relationship between species with the way we group manuscripts. Just as animals are divided into species, genuses, families, and so forth, we group manuscripts into families, text-types, and perhaps other groupings. In this particular case, the various sets of terminology in the different fields have actually informed each other, with Cladistics even starting to blur the lines between disciplines.
But, as Dawkins notes (The Ancestor's Tale, p. 399), of all these various levels of biological kinship, only one -- the species -- is rigorously defined: If two creatures interbreed, they belong to a single species. If they don't interbreed, they aren't. (Even this gets a little complicated when dealing with creatures such as bacteria which reproduce asexually, and there are other complications involving species which sometimes hybridize. But it's a workable starting point.)
The situation in textual criticism is similar: The only certain relationships are those involving immediate kin. Parent and child manuscripts are generally pretty clearly defined. (I wish I could say the same of sister manuscripts; they certainly should be clearly defined. But, having seen manuscripts which clearly are not sisters called sisters, we can't trust some declarations about that level of kinship.)
Just as any biological relationship other than the species is imperfectly defined, so a textual relationship beyond the immediate kinship level is somewhat arbitrary. Does this, then, mean that there are no other relations?
This can hardly be the case. The exact moment at which species split, and split again, and split again is uncertain -- such evidence as we have indicates that it is not a rigid dividing line, such that beforehand you have homo erectus and afterward homo sapiens, or beforehand a generic cat-like creature and afterward a lion. The distinction is clear now only because all the intermediate creatures -- the semi-lions and the intermediate-between-homo-erectus-and-sapiens have vanished. (Which adds to the confusion regarding manuscripts, since manuscripts of all ages survive. Biologists have only a few fossils. We have fewer manuscripts than we like, but it seems clear we have a higher fraction of old manuscripts than biologists have of the surely millions of members of species such as homo habilis or homo erectus. What's more, many of the creatures whose fossils have survived died young, without offspring, whereas the extant manuscripts may well have been copied.) But, with regard to manuscripts, we know that every one was copied from some number of source manuscripts and corrected from some number of others. We are guaranteed that splits occur, though we don't know when and at which point. The trick is to define the splits. Just as species did not instantly convert from one form to another, you probably can't say that a particular manuscript is of some type and a copy of it is of some other type. And, just as some species are closer to each other than others, so may one text-type be closer to another.
Dawkins, in fact, has a warning from biology about this, which surely applies in textual criticism as well: "Members of the cladistic school of taxonomists can become positively evangelical... [in] proclaiming the non-specialiness of fossils.... They take the sensible statement, 'It is unlikely that any particular fossil is the ancestor of any surviving species,' and interpret it to mean, 'There never were any ancestors!' Obviously this book stops short of such an absurdity. At every single moment of history there must have been at least one human ancestor... even if any particular fossil almost certainly isn't it." Similarly, in textual terms, every manuscript traces back to the autograph through similar manuscripts, even if those manuscripts haven't survived. L in the gospels probably is not descended directly from B, and B is probably not descended from P75, but L certainly has an ancestor written about the same time as B which was much like B, and B has an ancestor that was much like P75. Manuscripts with similar texts will have links at least somewhat more recent than the autograph.
Colwell once said that "Weak members of a Text-type may contain no more of the total content of a text-type than strong members of some other text-type may contain" (Studies in Methodology, page 33). This entirely fits what we learn from evolution. If I read Dawkins correctly, for instance, it appears that rabbits and mice split before humans and lemurs. Yet, based on the external characteristics at least, it appears that rabbits and mice are more similar than humans and lemurs. Humans and lemurs are more closely akin in terms of branching -- but the level of evolution along the primate branches has been extreme, whereas the rodents and lagomorphs (rabbits) have evolved more by specialization than by creation of new characteristics.
In this context, we note that degree of evolution is not the same as degree of kinship. Dawkins, (The Ancestor's Tale, p. 323), prints an evolutionary tree (phylogram) showing degrees of evolutionary divergence from the ancestor. I'm going to print a very simplified (and probably not very accurate) version of this as a demonstration. The longer a particular line, the more change in the creatures since the branching point.
|------------------- human
|-|
| |---------------- blue whale
|-|
| | |------------------ opossum
| |-|
| |-------------------------- platypus
|
| |------- birds
| |
|--|---|------ turtles and skinks
| | |
| | |---------- alligators
|-| |
| | |----------------- amphibians
| |
| |------------- lungfish
|
| |--- trout and salmon
| |---|
| | |---------- cod
|-|
|
|------ carp and loach
Note what this says: Although mammals such as humans and whales have undergone a much higher rate of evolution than trout or cod or carp, we are not descended from trout or cod or carp -- indeed, we are less related to them than to the surviving lungfish, even though lungfish are still pretty much fish and we are, well, us. All of the species involved have changed. It's just that we've changed more obviously. But we're all equally separated from the common ancestor. (Indeed, the fish are probably more separate in terms of generations; a fish generation is typically one year or less, whereas a hominid generation is many years.) Then there is the platypus: It's usually regarded as "primitive," and certainly it has primitive features such as laying eggs and secreting milk through the skin rather than having distinct nipples. But this doesn't mean it hasn't evolved. To survive with these primitive features, it has in fact evolved very heavily, developing (for instance) an electrical sense in its bill that allows it to detect small prey with its eyes closed. To get from the earliest mammals to human beings involves few "inventions" (we have a big brain, and hands -- but all mammals at least have some brains, and forelimbs). To get from early mammals to a platypus involves a new invention: That electrical beak. To develop that requires a lot of genetic change.
This again goes back to Colwell's point: Just as we are more related to birds than to cod, even though the differences between humans and birds are in many ways more dramatic than the differences between humans and fish, so the variations within an early text-type may seem nearly as dramatic as the differences between an early and a late text-type.
There is an interesting side point here, not experimentally verified but with some good mathematics about it, based on the work of E. Mayr. Mayr examined the process of speciation -- what happens when one species becomes two. It is widely believed that this usually happens because of some "separation event." A "separation event" comes about when some outside factor causes a population to split in two -- an example might be a river changing its course and dividing what, until then, had been a local population of some sort of mice, say, which cannot swim across the river.
As long as the members of the species could interbreed, it was hard to separate into two species, because everybody could breed with everybody else. New genes would tend to be swamped.
Divide the species in two, and there is no longer any interbreeding. The two halves will start to evolve separately, and so may more easily become separate species. This is not certain, note, but the separation makes it easier.
Thus far is generally agreed. Mayr's work comes in looking at the two populations. The conclusion is that the smaller population is more likely to change dramatically. This is simple mathematics: If a small mutation occurs, it is much more likely to survive in a small gene pool where it is less likely to be overwhelmed. The smaller the population, the faster new genes can spread -- plus it's quite possible that a smaller group will be subjected to more selection pressure.
When I read about this, I instantly thought of the local texts idea and their evolution. If you have a lot of manuscripts, change will be slow: There are always many manuscripts around to cross-check against. If a manuscript contains an error, it can be compared against another and corrected.
Now imagine a manuscript in an isolated monastery or some such place, with few other manuscripts to compare against. Perhaps they started with only one (or perhaps one of each section, or only one of the Apocalypse), and it not well copied. Having no other manuscript to be compared against, its errors will either survive or be fixed by conjectural emendation. As copies are made, additional errors will creep in. And, if the original was a papyrus manuscript, it will likely wear out in short order and no longer be available for comparison. Where there are fewer manuscripts, there may well be more diversity.
Now consider the Byzantine text: It is the most common text-type, and very unified. The Alexandrian text is smaller and less unified. The "Western" text is smaller still and even more diverse -- so diverse that we can't even agree on its boundaries. Is it a wild text -- or was it simply never widely copied? Mayr's theory implies it might be either.
This also brings up the constant question of "lumping" or "splitting." These have now become rather standard terms, with "lumpers" being biologists who place similar but not identical specimens in a single species, while "splitters" divide anything not shown to be identical. This can get pretty strange even for existing species -- in my lifetime, two types of birds, the Baltimore and Bullock's Orioles, initially listed as separate species, were lumped into one species, the "Northern Oriole," then split back into separate species again. And these are living creatures where we can observe their breeding habits! In the case of fossils, splitting can become so extreme that just about everything becomes its own species.
And species is the one definable term in biology. Splitting or lumping a family or phylum is certainly more arbitrary. Similarly, splitting or lumping text-types has to be arbitrary. The obvious defining point would be the last common ancestor -- but without having that common ancestor in hand, we don't have much to go on, we are forced to try to reconstruct. And that's with even if we ignore mixture.
The splitting/lumping issue is a significant problem. Suppose that the Byzantine prioritists are right and the Kx group of manuscripts in fact represents the original text. In that case, the Family P group of manuscripts probably could be considered a separate text-type. But if the Byzantine type is not original, then the P manuscripts are at best a sub-text-type.
This reinforces a point made above with regard to text-types. Evolutionary phyla do not have to be equidistant from each other. Some are closer than others. There has to be some level of difference, but there can be more than the minimum -- and the minimum can be relatively small. Rigid definitions are a mixed blessing: They allow us to speak precisely, but they must not be allowed to bind. New data must allow us to change our definitions, just as biologists have changed the genus and family classifications of many species over the years, and as astronomers recently (and quite correctly) downgraded Pluto from a planet to a dwarf planet. (They did so correctly because there is a difference in kind between Pluto and the other planets: Pluto is a typical Kuiper Belt object; there are lots of things in the same general area much like it. There is nothing like Jupiter in Jupiter's orbit.) The comparison to the Colwell-Tune 70% criterion for differentiating text-types should be obvious: The 70% difference plus gap was for the Alexandrian and Byzantine texts. There is no particular evidence that it applies to anything else. It's just as if you said that, because mammals and reptiles differ in 47 particulars (or whatever number you produce), then reptiles and birds must differ in exactly 47 or they can't be separate groupings.
This problem even infects the mathematical models of biological stemma. This is known as "long branch assimilation." If two particular species diverge far from the main bulk of specimens, parsimony analysis can pull them together. (Genetic trees with branches of this type are said to fall within the "Felsenstein Zone.") This certainly reminds me of the Claremont Profile Method and its infamous lumping of Codex Bezae with the Alexandrian Text. Keeping in mind also Colwell's warning that strong members of distinct text-type may share more readings than strong and weak members of the same type, might there not be other unnoticed examples of the same thing? (For a slightly artificial example of how this might actually come about with a text, see the appendix on The Bédier Problem in the article on Non-Biblical Criticism).
The key point here is how we use text-types. If they have any use at all, it is to supply relatively independent paths back to the archetype. This obviously argues strongly for the "ancestral" model of a text-type rather than the statistical model used by followers of the Colwell-Tune definition.
One place where genetics has curious traits is with regard to mixture. Generally when a lineage splits, that's it -- there can be no recombination. Dawkins in The Blind Watchmaker, p. 248, notes for instance that humans have eight distinct genes for making globins (hæmoglobin, etc.) These genes are all believed to be descended from a single original globin gene, even though they now exist on separate chromosomes. Because they are separate genes, in separate places, they can no longer mix; we can create a stemma and perhaps recreate the original globin. But if we did, it couldn't be used for anything; the current globins are now separate and individual. At least in theory. It's largely true in practice, too, for animals, since they only mate with their own kind. It's a lot more complicated with plants, where foreign pollen can sometimes show up. I have, frankly, no idea how this affects things; it might be worthwhile to find out.
While we're talking species, and text-types, and mixture, we might also mention the curious phenomenon of "ring species." This is an unusual phenomenon because it requires a sort of habitat loop -- a region of territory where animals can live at the boundary but cannot cross the middle. An example is the shores of a very large lake (either in or out of the water), or a particular elevation in a valley surrounded on all sides by mountains. Dawkins (The Ancestor's Tale, p. 301) offers the example of California's Central Valley and the Ensatina salamanders. These salamanders live only at a certain elevation. Since the valley is entirely enclosed, their habitat looks like a very elongated letter "O," about four times as tall as it is wide, with a very large region in the middle where the salamanders cannot live. If you start in the southwest corner of the loop, you'll find salamanders with plain skins. Move north along the western side of the valley, and you'll gradually see slight blotches appear on their skins. Once you reach the north end, turn south, and travel along the east side of the valley, you'll find that the salamanders' skins will get more and more blotchy; eventually, when you get to the southeast corner of the ring, they are extremely blotchy.
Now here is the interesting point. Start at any point along the ring, except the southernmost part, and salamanders will breed with their neighbours to the clockwise and counterclockwise directions. There is no species distinction. But, down at the southern part, you have plain-skinned and blotchy-skinned salamanders, and they will not interbreed. In other words, there is a species distinction there, even though there is continuous variation around the circumference of the ring. (Anti-evolutionists claim we've never seen a new species created in the lab. The ring species, however, is a pretty good example of how it comes about. If a flood or earthquake or human activity were to destroy the northern end of the Central Valley, this continuous species would suddenly become two species! Thus we could do it if we wanted to.)
Ring species probably can come about in either of two ways. One is to have an intermediate species split in two (in the example above, a slightly blotchy species of salamander might have arrived at the northern part of the central valley and spread in two different directions); the other is to start with one distinct type and have it slowly crawl "around the circle" (in the above case, the ancestor was either a very plain salamander in the southwest or a very blotchy one in the southeast, which bred only one way around the circle). Could textual "mixture" also arise by both means? That is, could one "mixed" manuscript be the ancestor of two distinct types of text (a common original splitting in two because of genetic drift), while another derives from combining two types of text (a hybrid)? There is no logical reason why not. The implications, however, are very different. In the former case of a split, both types have value (since they lead back to an earlier archetype), and their agreements have particularly high value. In the case of the hybrid, though, the hybrid text has no value unless the ancestral texts are lost. (This is, of course, precisely what Hort said about the Byzantine text, and why he denigrated it. And no one has seriously contested the logic; the question is whether his history of the text is accurate.)
The example above also points up the generally-agreed cause of separation into species: Sudden (usually unexpected) geographical separation. If a lake is split in two by an earthquake, say, fish in the two halves, originally one species, may evolve into two. Indeed, Dawkins (The Ancestor's Tale, p. 341) observes that, for some fish species, a gap of just two kilometers between reefs in a lake is enough to cause genetic separation. Is this not much the same as the proposed origin of local texts, where a particular region gradually standardizes on a peculiar text?
Evolution may also say something about the nature of variants. You've probably heard that most creatures' DNA is rather a jumble of stuff. It isn't as if someone set it up in an intelligent, orderly manner; it's scattered all over the place on the chromosomes. (Just about what you would expect of stuff that has evolved and changed and developed new purposes over the years.) Much of it, in fact, no longer has any purpose at all, and is not used. (This might, for instance, include DNA for making gills in humans: we don't live underwater any more, so we don't need gills -- and even if we had them, as warm-blooded creatures we can't derive enough oxygen from sea water to meet our needs.) This is the so-called "junk DNA" -- a name which accurately describes how useful it is, though a better historical name might be "fossil" or "discarded" DNA.
The interesting thing is, this unexpressed DNA tends to change faster than DNA that actually does something. This makes sense, when you think about it. If a gene needed to create, say, the human lung mutates, the probability is high that the mutation will be detrimental -- quite possibly fatal. It will probably die out. But a mutation that affects the no-longer-used genes that make gills or whatever can be preserved, because the changes have no effect on the survival of a human who doesn't use gills anyway.
This at least bears thinking about. There are only two significant many-verse variants in the New Testament: Mark 16:9-20 and the story of the Adulteress. There are more than that many add/omit article variants in the average chapter. You might argue that the latter is a meaningful variant -- but it isn't very meaningful. In the case of a definite article before a name, it means almost nothing -- and these are incredibly common. (If you check the section on Most Uncertain Readings, examining just the Gospel of John, we find two dozen variants ihsous/o ihsous. That's not variants in one or two minor manuscripts, note; they are variants substantial enough to cause real differences between editors.) Could it be that minor variants will survive better than major? Of course, that needs to be tested -- but there are at least a few manuscripts that seem to bear this out. A in the Gospels is mostly Byzantine -- but it has a much higher number of major Alexandrian variants than lesser Alexandrian variants, as if its ancestor were a Byzantine manuscript loosely corrected against an Alexandrian manuscript, but with only the major differences noted.
There is a curious analogy to this in some a sexually reproducing species. Most species of rotifers reproduce sexually, the equivalent in a text of mixture. But there is a group that reproduces asexually -- no mixture, just direct copying. It seems pretty clear that they are descended from an ancestral species that reproduced sexually, since their chromosomes still come in what superficially appear to be pairs (five pairs, ten chromosomes total).
Now here is the interesting point. These bdelloid rotifers are now regarded as having divided into 360 species. All derive from one ancestor and her ten chromosomes. All have undergone some mutation and evolution. But, because the chromosomes no longer mix and match, the chromosomes of each pair are free to evolve away from one another. There is no longer a need for chromosome 4-left to be interchangeable with 4-right. And indeed, they no longer are. What were once five pairs of chromosomes are now, in effect, ten single chromosomes.
And it gets more interesting than that: Often the chromosomes display less difference between species than within species. That is, if you compare 4-left in species A, it is more like 4-left in species B than it is like 4-right in species A. Indeed, there may be more variation in 4-left within species A than there is between A's 4-left and B's 4-left. Here the analogy is perhaps to text-types; if we cannot know how much internal variation there is within a species, does that not imply that we cannot know how much internal variation is in a text-type?
A sidelight on this is provided by the human X and Y chromosomes, the only unpaired chromosomes in the genome. These chromosomes have developed what are called "sexually antagonistic genes." The X and Y chromosomes, being distinct, no longer need to share genes, and in fact can develop genes which suppress each others' actions. This, in fact, is pretty much what it means to be male: A handful of genes on the Y chromosome have suppressed the process of becoming female. The fact of sexually antagonistic genes, in fact, is believed to be why the human Y chromosome is so small: In the population as a whole, it's outnumbered 3:1 by the X chromosome, meaning that X can evolve much more quickly. So where the two conflict, Y is going to lose. As a result, it abandons the battle; there are few genes still in use on Y. Small as it is, it's mostly junk DNA. Most of the genes that used to be on Y are believed to have moved to other chromosomes.
This antagonism can be extraordinarily real. This has been shown in fruit flies. The seminal fluid of the male has been shown to try to increase female ovulation and suppress the sexual urge -- in other words, the male fly uses its semen to try to make sure the female doesn't mate with anyone else. The female genes, naturally, want to keep their choices open, so they evolve immunity to the chemicals in the semen.
At least, in nature they do. Matt Ridley, in Genome, pp. 113-114, describes an experiment undertaken by William Rice, in which a population of male flies were allowed to keep evolving more aggressive semen, while a population of females was made to keep its old semen resistance genes. After 29 generations, Rice reunited the breeds -- and the male semen had grown so strong as to be irresistable, even fatal, to the unevolved females. (This, incidentally, should pretty well answer any questions about evolution not creating new species. Rice's flies are already on the brink of speciation; another few dozen generations of that and the old and new flies functionally couldn't interbreed at all -- the males might try, but since the females would produce no eggs, the matings would be infertile.)
This sort of thing probably can't happen in the Bible text, because mixture does not involve the same sort of antagonism. (Also, the Bible doesn't breed as fast. There is some mathematical work on how fast a population can be traced back to a common ancestor; Dawkins summarizes it on pages 42-45 of The Ancestor's Tale. It involves many assumptions, and we have to add more to apply it to the text, but as a general model, it appears there are only about fifteen generations between the manuscripts surviving today and their primary archetype, and only seven to ten between that archetype and the autograph. This isn't enough generations to allow speciation except in the most extraordinary circumstances.) But it bears some thinking about. When text-types mix, which readings will tend to be perpetuated? Answer: It appears, in general, the longer ones; scribes didn't want to risk leaving out any words that might be original. There are exceptions (424 being the obvious example). But they do appear to be the exceptions.
Note the implication: Because the Byzantine text is the longer, fuller text, its readings will tend to prevail at any stage of mixture. Does this mean that this is how the Byzantine text came to prevail? No. What it does mean is that, all else being equal, the Byzantine text could be expected to prevail.
So much for "normal transmission." It applies, at best, only where there is no mixture.
Some footnotes. There seems to be a common belief that evolution has some sort of appointed end (typically us). This is an extreme and dangerous misconception. Evolution always operates after the fact: First variations arise in the population, then natural selection acts on them. To give an analogy: Suppose you're on the way somewhere (say a shopping mall) and you come to a branch in the road. From there, there are two ways to get to the mall, depending on whether you take the left or right fork. Once you take the a particular fork, your route is pretty well determined. But until you make that decision, it's entirely up in the air.
Similarly, many evolutionary problems have multiple solutions. As an example, consider the disease rickets, caused by an inadequate supply of Vitamin D. This was not a problem for the dark-skinned early humans, who lived in Africa; they didn't need to cover their skin for warmth very often, and they got full sunlight all year long.
Take someone like that to Scandinavia and he or she might well die of vitamin D deficiency. (Or so I heard many years ago; I don't know if this analysis has held up in the days of DNA analysis. In any case, the analogy is correct.)
There are several possible solutions to this problem. The body could evolve to manufacture vitamin D some other way. Or it could discover some alternate source (this is what the Inuit did, for instance -- they get their vitamins from the organ meat of seals, and remain relatively dark-skinned). Or -- Scandinavians could develop light skin, allowing sunlight to create Vitamin D more easily. All three of these techniques are equally valid (at least for the vitamin D problem). But Europeans ended up with #3, while the Inuit use #2.
Evolution is always like that. It does not necessarily find the best solution to a problem; it finds better solutions than what went before. A species suffering too much predation may learn to run faster, or develop better camouflage -- or it might just start breeding faster to keep the population up. Do this long enough, and you might get a very superior creature of some sort -- but it can take a very long time, and the outcome can be quite unexpected. Indeed, it can be a very poor adaption for any circumstances but its own; this is why it was so easy to drive so many species, such as dodos, extinct. An example of an adaption that works only in its peculiar context is the cave fish that have lost their eyes: In pitch darkness, eyes don't help and leave them with a vulnerable spot. Occasionally, of course, such fish get washed out of the cave and into daylight, where they find themselves at a huge disadvantage. That being the case, the best solution for the cave fish would not have been to lose their eyes but to develop some sort of very strong eyelid to protect it. But, for the fish still in the cave, this solution is no better than losing their sight, and probably harder to develop. So the fish go blind because it's a workable answer to a genuine problem. Such "quick fixes" are found in the human genome, too: we've already mentioned the sickle cell mutation. This seems to have been quick-fix human answer to malaria, which became more common when human clear-cut land, making more breeding sites available for malaria mosquitoes. A better solution would have been to develop real immunity to malaria, but sickle cell was better than nothing, and it was an easy mutation to create. Similarly, the cystic fibrosis gene is effectively fatal to people who have two copies -- but people with one copy seem to be nearly immune to typhoid (see Kevin Davis, Cracking the Genome, p. 47). So that gene, which is as dreadful as the sickle cell gene, is common in Europe.
This is perhaps a useful warning for the textual critics who have tried to reduce scribes to automata. A scribe confronted with what appears to be an error may try to fix it -- but you probably can't predict the fix. If it's a "good" fix (according to whatever definition of "good" other scribes use), it may well propagate. But you can't predict the fix before it's made.
Another interesting point about evolution is the modern concept of "punctuated equilibrium." Evolution is not continuous -- when conditions are stable, evolution operates very slowly, with few changes over the years. Upset the stable conditions -- due to climate change, or the arrival of a new species from outside, or just a wildly successful mutation -- and the whole thing has to, in effect, scramble to find a new equilibrium. These are the conditions under which species more rapidly go extinct and new species are created.
Now think, for instance, of the various persecutions of Christianity. Emperors such as Diocletian destroyed every Bible they could get their hands on, and took out a certain number of scribes as well. This means that there is little opportunity to compare manuscripts, and many of them will be copied secretly and by amateur scribes. This will probably encourage "mutations" -- new variants and text-types evolving. We can't see this happen very much (the persecutions and later barbarian invasions took place at a time when the manuscript record is very thin), but it seems reasonable that this might happen.
While we're looking at history, let's take one note from another evolutionary process: That of language. We mentioned above the Proto-Indo-European language that is the ancestor of (among others) English, German, Latin, and Greek. Since we have a stemma of sorts for this language's descendants, we can reconstruct most of the language (basically by taking the grammar of Sanskrit and reconstructing the vocabulary by stemmatic principles). This reveals interesting points. For example, the reconstructed language has a number of words pertaining to agriculture. This implies that it originated after agriculture had spread to their regions. (It has been hypothesized that the success of the Indo-Europeans was due to their possession of agriculture). Certain other words -- e.g. relating to pastoralism and to metals -- are not found, implying that it predates the introduction of those habits.
This sort of conclusion based on linguistic data is pretty fragile, especially as regards the argument from silence. (It's theoretically possible, e.g., that the Indo-Europeans herded, say, gerbils, but because none of the societies descended from them did gerbil-herding, all relevant words died out and were reinvented later.) But sometimes they can be important. There are occasional analogies to histories of texts. I can't think of a good one for the New Testament, but consider Isaiah 7:14. If a version translated from the Hebrew says that a young woman shall bear a son (which is, of course, the correct reading of Isaiah), it tells us very little. But if a version translated from the Hebrew says that at virgin shall bear said son -- a misrendering which originated with the Septuagint -- there is probably Christian influence in there somewhere. Or at least LXX influence.
One key aspect of evolution has little relevance to Biblical texts: The so-called "arms race." This is what comes about when two species are in particularly close competition -- say an herbivore and a predator, where the predator is the primary killer of the herbivore and the herbivore is the predator's primary food. Call them, for the sake of simplicity, antelope and leopard. They can't escape each other (barring a habitat change or the intervention of another species). If antelope start running faster, then leopards will get faster in response. If antelope learn to hide more effectively (by disguising their appearance or scent), then leopards will develop better eyes or ears. If antelope start to breed faster, then more leopards will be able to survive to consume the larger population. No matter what one species does, the other will find an answer (or go extinct). As the years pass, they have to devote more and more energy to leg muscles, or to fancy fur patterns, or extra offspring, or whatever it takes to survive. If there were a way to call the whole thing off -- for the antelope just to give in and say, "Here. Take ten percent of us each year (or whatever percent the leopards actually take) and stop evolving," it would make life easier for both species. But they can't; they're stuck. Ultimately, this is very like the mathematical problem known as the "prisoner's dilemma" (see the article on Game Theory).
This, as noted, does not occur in textual criticism -- though it does sometimes seem to occur in denominations, as preachers desperate to build congregarions threaten worse and worse hellfire. I do sometimes wonder about a sort of a race between plain-text manuscripts and lectionaries -- since lectionaries were more useful in churches, there would be pressure on plain-text manuscripts to become more and more useful for church reading, so that lectionary incipits and such would be put into the margin -- and even into the text (as happened, e.g., with 1799). I doubt this has affected manuscripts seriously -- though it's had some pretty strange effects on English translations. (I have a New Revised Standard Version with a marginalia designed, I think, for a King James Version by a very conservative scholar. At times, it seems as if text and margin are at war; at others, the marginalia, by adding section headings and such, appear to me to at least distort the meaning of the text by dividing sections which should be united.)
Finally, a word about evolutionary progress versus evolutionary direction versus randomness. I've repeated several times above that evolution does not result in progress -- that, e.g., a mole is not "better" than, say, a weasel, at least in the general sense of surviving; they're just different. There is a tendency to think that, if a process doesn't have a destination, it must be random. This is not at all true. Evolution doesn't have a destination, but it is rarely random. Dawkins, in The Blind Watchmaker (starting around page 65), points out the difference between random selection and cumulative selection. This is a very important point. Random selection, in which you simply throw a bunch of traits together, has no direction and would almost never produce improvement. Cumulative selection is altogether different.
Let's take a very simple test (Dawkins does something like this, but I'm going to produce a variation which I think makes it clearer). Let's produce 10-digit strings of numbers, consisting of the digits 1-3. Call each 10-digit sequence a "creature." So 1111111111 is a creature, and 1231231231 is a creature. It can be shown that there are 59049 different creatures of this type.
Now here is our goal: We want to create a creature that has all digits belonging to one set. We don't care which set; just all belonging to the same set.
Remember that there are 59049 creatures. There are only three of them which meet our criteria: 1111111111, 2222222222, and 3333333333. So if you just set up a random creature generator and have it spit out creatures, only one creature in 19683 (59049 divided by three) will be of this type.
To demonstrate this point, let's try 25 generations of random selections. My trusty spreadsheet gave me this list:
3232232231
2123121131
2222221132
3223112123
1323112223
2223112312
1321331222
3223113112
1223333321
1123233333
2222133331
3322323323
2123322232
1332311311
2111132322
1133123223
1311113132
3121213322
2211223121
2313313223
2131132323
3223222222
2313121322
1112312211
1223213312
Not one of them is acceptable -- in face, only eight of them have even the first two digits the same!
That's random selection. Chance never gets us anywhere. If by some coincidence you get something close to an acceptable form -- well, it will all be jumbled in the next generation, so even being 90% right is no good. Under random selection, you have to have everything right the first time. Some people claim that this is so improbable that evolution can't work. And they may well be right -- except that random selection isn't how evolution operates.
Keep the problem of random selection in mind as we consider cumulative selection. Here's how cumulative selection works: Once you find a partial solution, you refine it and converge toward a better form of it. For example, take our first creature above: 3232232231. It has, by chance (or, perhaps, by defect of Apple Computer's built-in random number generator), only one case of 1, five of 2, and four of 3. So this creature "inclines toward" 2.
So our goal is to "evolve it" toward 2. Note that the choice of 2 is entirely arbitrary. If there had been more instances of 1 than of 2, we'd say our creature inclined toward 1, and evolve that way. But this one inclines toward 2. So what we do is, we hold every instance of 2 to be fixed, and let only the other digits evolve. How long does it take to get from 3232232231 to 2222222222?
Answer: In my first run, it took all of three generation:
3232232231
1222222223
1222222223
2222222222
Remember, in random selection, it would almost certainly have taken thousands of generations.
Of course, this is extreme. We started with a number that was very two-heavy. Let's try this a few more times. Remember that we will select for whatever is the most common number in our first try. Here are the results of six more tries:
|
1122311331
|
3233123222
| 1112332333
|
1331213223
|
3331213223
| 2311123132
|
This shows the absolutely astonishing power of cumulative natural selection. We have achieved our result about a thousand times faster than randomly assembling traits. And if we were trying to achieve something like a living being, with an even lower random probability, the advantage of cumulative selection would be even greater. (There appears to be a logarithmic relationship here, though I haven't seen math on this and am too lazy to work it out. Suffice it to say that cumulative selection produces a huge improvement in evolutionary speed.)
The example above may sound improbable -- shouldn't there be a best way for a species to survive? By no means. As we've noted several times, many survival problems have multiple solutions (e.g. better concealment, better eyesight to spot predators sooner, higher speed to outrun predators, or faster reproduction to have offspring before the predator can kill you). Sometimes the different "cures" can be just about very similar in style and nearly equal in effectiveness. Let's try one more example -- consider an arbitrary species of flowers (call them rolys -- sort of half way between a rose and a lily) and pollinators. It happens that bees cannot see the color red, so they will not pollinate red flowers. Hummingbirds, as a result, are specially attracted to red flowers, and pollinate them; if there are enough red flowers, they may ignore something with petals of another colour.
Now suppose a species pf rolys shows up where some of the flowers are red and some are blue. The red ones will be pollinated by hummingbirds, the blue by bees. A red flower, as a result, will never be pollinated by blue pollen, and and blue flower will never get red pollen. There are only two possible outcomes: Either cumulative selection will apply, and one or the other color will come to dominate (presumably red flowers if hummingbirds are more common, or blue if bees are the primary pollinator) -- or the species will split into two, with red rolys and blue rolys not interbreeding and eventually becoming separate species.
This could perfectly easily happen to literary events, too. One that occurs to me is the reporting of miracles. Christianity regards miracles as validating its truth. As a result, we see non-miraculous events treated as miracles (see, e.g., the story of Eutychus in Acts 20:9-12. Paul didn't do anything; he didn't heal the boy, and for all we know, Eutychus died later or was permanently crippled. But it's given the feeling of a miracle. Or consider Elisha and the Shunammite's son. 2 Kings 4:34 describes what sounds like mouth-to-mouth resuscitation -- but, again, it's treated as a miracle).
Islamic tradition is the reverse: Mohammed denied that he was a miracle-worker. I can't recall a single human-worked miracle in the Quran -- not that I'm expert. But I consulted the translation of Abdullah Yusuf Ali, which is carefully indexed. It led me to Surah 29:50, which notes the lack of miracles; "The Signs are indeed with God [and not human beings]." The rather indignant footnote complains about unbelievers asking for signs (beyond the what are regarded as the self-evident signs of the existence of the universe, life, and such): "Everything is possible for God, but God is not going to humor the follies of men or listen to their disingenuous demands He has sent an Apostle to esplain His Signs clearly, and to warn them of the consequences of rejection. Is it not enough?"
You don't have to believe in Christianity or Islam to see how extremely divergent these positions are! Clearly, if a slightly strange event happens, Christian folklore will tend to turn it into a miracle; Islamic folklore will have no such tendency.
Now forget folklore, and miracles, and think texts. Start with the same original text, then hand it to an orthodox Christian, an Arian (who considered the Son to be inferior to the Father), and a Nestorian or other monophysite who believed that Jesus was God and not man, or man and not God, or a figment of the imagination, or something like that. Without deliberate alteration, each scribe will probably make errors that support his theology. The resulting text will clearly be Orthodox, or Arian, or Monophysite -- and will encourage its copying by other scribes with that theological tendency, making the second generation even more partisan, and so on. It's very much like the cumulative selection case. Once the process starts, it will tend to proceed in the same direction because it's self-reinforcing.
Much of what has been said above is about evolution. I want to repeat something I mentioned above: Evolution today is not your parent's evolution, nor Darwin's. We are living in the era of genetic Darwinism -- neo-Darwinism.
By that I mean that the modern understanding of DNA has completely changed the
field. To see why, we probably need to say a few words about genes and genetics.
I won't burden you with much history. Suffice it to say that, by the mid-twentieth
century, it was clear that most of the chemical work in the cells was done by enzymes,
and enzymes were made of proteins, and that the data needed to assemble those
proteins was stored in DNA -- deoxyribonucleic acid.
 Despite the name, DNA is not a
single molecule, but rather an infinite class of molecules, built from five smaller
molecular components which in turn combine into four basic pieces of DNA. The five
molecules are of two types. Four -- the "nucleotides" --
are used to store information, while the fifth
provides the framework in which the other four are stored. The four
pieces can be thought of a T-shaped elements which assemble like a puzzle. The cross bar
of the T is a phosphorus-based "backbone" (this, incidentally, explains
why phosphorus spills at sea cause algae blooms: Phosphorus, since it is part of DNA,
is absolutely essential to all life -- and phosphorus is present in a much lower
proportion in sea water than in living creatures. The limiting factor on an ecosystem,
especially a water ecosystem, is almost always phosphorus. Add phosphorus and you
get a population explosion lasting until something else becomes the limiting factor.
Life is "accustomed" to phosphorus shortages; it is not "accustomed"
to phosphorus abundance, and goes out of control if the supply is large enough.)
Despite the name, DNA is not a
single molecule, but rather an infinite class of molecules, built from five smaller
molecular components which in turn combine into four basic pieces of DNA. The five
molecules are of two types. Four -- the "nucleotides" --
are used to store information, while the fifth
provides the framework in which the other four are stored. The four
pieces can be thought of a T-shaped elements which assemble like a puzzle. The cross bar
of the T is a phosphorus-based "backbone" (this, incidentally, explains
why phosphorus spills at sea cause algae blooms: Phosphorus, since it is part of DNA,
is absolutely essential to all life -- and phosphorus is present in a much lower
proportion in sea water than in living creatures. The limiting factor on an ecosystem,
especially a water ecosystem, is almost always phosphorus. Add phosphorus and you
get a population explosion lasting until something else becomes the limiting factor.
Life is "accustomed" to phosphorus shortages; it is not "accustomed"
to phosphorus abundance, and goes out of control if the supply is large enough.)
The stem of the T is where the real information is stored -- in an elaborate hexagonal or hexagonal-plus-pentagonal structure . These structures consist of one of the four nucleotides. The four are usually referred to by initials, A, C, G, and T; they are properly called adenine, cytosine, guanine, and thymine. (In RNA, which is a parallel to DNA with a somewhat different backbone, U=uracil replaces thymine.)
The result really is like a children's toy. There is the rigid brace of the phospate backbone, the structure of the nucleotide coming off of it, and a couple of spots where hydrogen bonds can hook, like the joins of tinker toys or the raised bumps on Lego blocks.
Note how beautifully functional all this is: Two of the nucleotides, A and G, are wide, with a double ring; the other two, C and T, are smaller. This means that the two possible links, A+T and C+G, are the same width. The one major difference is that the A/T linkage, which has three hydrogen bonds, is slightly stronger (and in fact is slightly more common in DNA; it has been speculated that this is so the DNA itself is more firmly held together).
That clever linkage, in which the A+T and C+G assemblies are the same size, means that DNA can form the famous "double helix," If we untwist a strand of DNA into a sort of ladder, it might look something like this:
Normally, of course, this is twisted around itself -- hence the "double helix" description (actually only a single helix, merely one with a twisted ladder-like form rather than a straight strand, but let's not worry about that).
Since there are four and only four possible nucleotides as we read along the DNA strand, it will be evident that this is a digital code, done in base four.
Well, theoretically base four. In fact, like Hebrew, "words" are grouped in blocks of three letters. That means there are 64 possible "words."
In a curious feature of the code, which has its advantages and disadvantages, all of these 64 words have meanings, even though there the number of messages is smaller than 64. That is, there are only so many messages a word has to convey. Possible meanings are one of the twenty amino acids used to make proteins, plus a few control commands (e.g. there needs to be something to say "stop" or "this is the end of the protein.") So the sequence AAA might stand for the amino acid alanine, AAC for argenine, AAG for asparagine, etc., with a bunch of sequences meaning nothing at all.
It doesn't work that way. Every one of the 64 possible words stands for something, meaning that there are about three ways to encode for each of the standard commands. (In fact some amino acids are represented by as many as six different codes, others by just one.)
One noteworthy element of the system is that all creatures tested use the same DNA code. This is, in one sense, astonishing -- one DNA code is as good as any other in terms of information storage. So if all species were independently created, they would be expected to use different codes. But once a code is established, it's effectively impossible to change it; the result is chaos. So the fact that all known creatures use the same code strongly implies common origin.
The obvious disadvantage of the all-codes-have-meaning system is that it makes it hard to detect damage -- if we had only 21 codes (say, arbitrarily, AAA through CCA, or perhaps some other scheme where the codes are spread throughout the alphabet), then if AAA mutated into GAA, we could be sure there was a mutation and set about trying to fix it. (Francis Crick, the co-discoverer of the double helix, in fact proposed an encoding scheme with a very strong error-correcting element in it: it made it impossible to misread DNA. Unfortunately, the language of DNA is not intelligently designed, and doesn't follow Crick's encoding pattern.) The side effect of this system with no error correcting is that a lot of genetic diseases can crop up by random mutation at a single nucleotide -- as Queen Victoria of England apparently suffered a mutation which gave her one gene for hemophilia. She herself, having a good gene to cover for it, did not suffer hemophilia. But many of her descendants did -- including, famously, the son of Nicolas II of Russia, with truly disastrous consequences.
The other side of the coin is, this system does make mutations easier. Most mutations are deleterious, as in Queen Victoria's case -- but some are advantageous. Apparently bad mutations are not too high a price to pay, as long as there are occasional good mutations, too. (This actually makes some sense: Most individuals in most species don't live long enough to breed, so losing a few sick individuals to bad mutations costs little, while even a few advantageous mutations can have a high payoff. It is, by ordinary standards, very cruel -- but it makes sense, evolutionarily.)
But this makes geneticists practice a discipline almost like textual critics. DNA is a series of words -- words which are all the same length, and with a total vocabulary of only 64 words, but they are words. And they form sentences -- complete genes, which encode complete proteins. And the sentences, unlike the words, can be almost infinitely long and complex.
Consider the implications. Suppose we compare the genes for making, say, hair in five different species. I'm just making this up to demonstrate the point, but the principle holds. Suppose the hair gene looks like this in our five species:
WORD: 1 2 3 4 5 6 7 8 9 10 chimp: AAA * GAC * GAG * CAG * GAA * TAA * TAG * GAT * CCC * CAG ... hippo: AAA * GAC * CGG * CAG * GGA * TTA * TAG * GGT * CCA * CAG ... human: AAA * GTC * GAG * CAG * GAA * TAA * TAG * GAT * CCA * CAG ... mouse: AAA * GAC * GGG * CAG * GAA * TTA * TAG * GAT * CCA * CTG ... whale: AAA * GAC * CGG * CAG * GGA * TTA * TAG * GAT * CCA * CAG ...
Now here is the part that resembles textual criticism: We can and should actually prepare a collation of this data. In this very simple example, we can do this by listing which species have which coding for each word. That's this (we should note that I've create a very high rate of variation. It's quite likely that, for any given gene found in all five species, there will be less than one variant in every ten words, rather than the seven per ten words I've illustrated). I've listed the majority reading first in all cases, taking humans as the standard in the event of a tie.
Word 1: AAA (all species)
Word 2: GAC (chimp, hippo, mouse, whale) ] GTC (human)
Word 3: GAG (chimp, human) ] GGG (mouse) | CGG (hippo, whale)
Word 4: CAG (all species)
Word 5: GAA (chimp, human, mouse) ] GGA (hippo, whale)
Word 6: TTA (hippo, mouse, whale) ] TAA (chimp, human)
Word 7: TAG (all species)
Word 8: GAT (chimp, human, mouse, whale) ] GGT (hippo)
Word 9: CCA (hippo, human, mouse, whale) ] CCC (chimp)
Word 10: CAG (chimp, hippo, human, whale) ] CTG (mouse)
As noted, we have seven variants. Four of these (words 2, 8, 9, 10) consist of singular variants. That leaves three variants we can consider meaningful:
Word 3: GAG (chimp, human) ] GGG (mouse) | CGG (hippo, whale)
Word 5: GAA (chimp, human, mouse) ] GGA (hippo, whale)
Word 6: TAA (chimp, human) ] TTA (hippo, mouse, whale)
Note that, in these variants, chimp and human agree every time except for the singular reading at word two (we might wildly guess that this is the gene that allows humans to grow long hair; nearly all other mammals, including chimps, have genes which give them a fixed hair length), as do hippo and whale, with mouse agreeing once with chimp/human, and once with hippo/whale. Based on this evidence, the genealogy for these creatures is:
proto-mammal
|
--------------------------------
| | |
proto-ape proto-mouse proto-water mammal
| | |
-------- | --------
| | | | |
chimp human mouse hippo whale
This is, of course, far too little evidence to go on, and the entire example is faked -- but the principle is perfectly valid, and shows how closely parallel genetic analysis is to stemmatics.
There is another note here. It's one thing to have a gene, and another to use it. We mentioned above that genes are like a digital data store. They resemble computers in other ways, too: They resemble computer subroutines, and the whole thing is a vast computer program. Like a computer subroutine, the same gene can be used by other genes for various purposes (there are actually molecules to turn a gene on or off, and many genes respond to multiple different activating molecules. Indeed, they may produce different proteins based on different activators). The whole system is very much like passing parameters to a subroutine.
This also explains why some cells are different. Many programs are customizable -- every word processor I've encountered in recent years has some sort of "work" menu on which you can stash commands you use a lot. Cells are like that, too: Very early on in the life of the organism, they get a chemical cue which activates a few master genes, and those master genes then tell them to become heart or liver or skin cells. And, in the very early stages of development, many of those cues come from chemicals passed on by the mother or the father -- a phenomenon known as imprinting. This is a relatively recent discovery, and I can't go into it deeply (I don't know enough, and the field is moving so fast that whatever I could write would soon be out of date anyway). But this gives us one more analogy to scribal copying -- a scribe might well be given instructions by his superiors to incorporate this, that, or the other set of marginalia, or to copy in uncials or minuscules, or any of a dozen other things. The basic text -- the "genes" of the manuscript -- may be unchanged even while looking very different and servinga very different function.
The statement at the top of this page about scientific theories of creation -- that the Big Bang, evolution, etc. are compatible with the Bible -- probably isn't going to convince anyone who isn't convinced, but on the off chance that there is a person out there who actually wants to give the matter a hearing, I'm going to offer further justification. Pointless, I know -- but hey, voting is pretty pointless (since I'm outnumbered two hundred million to one), and I still vote anyway; I just never see anyone worthwhile elected. So I'll spend a little time tilting at this windmill.
I'm not even going to address Intelligent Design, since it is a farce posing as science -- and nobody believes it anyway. This is addressed solely to the issue of whether there is a fundamental conflict between Genesis and scientific theories of creation (as they are understood early in the third millenium of the common era; there is of course every expectation that they will change. Scientific alternatives have long been offered to the Big Bang. Personally, I've never been very fond of the Big Bang -- it's rather a lot of theorizing based on very little data. But it's currently the accepted model of universal formation, and it's certainly the one most like Genesis. In any case, eliminating the Big Bang doesn't eliminate the stumbling block of evolution; every scientific model of the universe assumes evolution as the source of biodiversity on earth.)
I would argue as follows. The current "inflation" model of the Big Bang very closely resembles Genesis -- e.g. in the early moments of creation, everything was a big jumble. There is no better description than "without form and void." The whole thing was a plasma. Then the three linked forces, strong nuclear, electromagnetic, and weak nuclear, split, "and there was light."
Gradually the whole mess settled down, and then life started to appear. It wasn't all at once, but nowhere does Genesis say bacteria and frogs and lions were created at the same time, merely that they were created at the same stage of creation -- after the creation of the universe, and before humanity. Humanity came last -- and, indeed, we are one of the last evolutionary results of one of the most recent branches of the evolutionary tree.
There are other interesting places where evolutionary theory can explain some curious aspects of the Bible. For example, why is it that the lifespans of the Patriarchs in Genesis keep getting shorter? It must be remembered that evolution doesn't care what happens to a living thing after it breeds. Mayflies and Pacific Salmon breed and die -- they don't even try to survive after laying their eggs. So there is no evolutionary reason not to experience senescence if you can breed well before that.
And mutations can cause senescence. The vast majority of mutations are detrimental -- e.g. cystic fibrosis, which is caused by a single change in a single gene, kills people young, and in most circumstances is purely detrimental. These mutations die out (cystic fibrosis still exists because it's a recessive gene and because it can happen by new hot spot mutations).
But there are mutations which can have mixed effects -- suppose, e.g., that human females once could have only three children. If a mutation came along that allowed them to have twenty children, but caused them to grow old faster, well, from the standpoint of evolution, that's a good gene. Chances are that aging is in some way a side effect of mutations like this, and their evolutionary consequences. (In fact, I just heard a very brief item about a gene that is known to do exactly that: The gene prevents cancer but accelerates aging. In a situation where few individuals live long enough to die of old age anyway, the cancer-preventing effects are doubtless worth the cost.)
I admit that it's a long, long stretch from this to the Patriarchs, but it's an interesting case of Genesis paralleling something that really does seem to have happened. It is particularly noteworthy that the steep decline in lifespan came in the generations after Noah -- noteworthy because the Flood would have caused a sharp constriction in the gene pool (something that geneticists have long thought to be true because the human genome has so few variations, something which could imply that, at one point, the population shrank almost to vanishing. The disaster involved may not have been a flood, but there was something.) A small population, subject to genetic drift, could easily lose certain genes, including those which promote longevity if they affect fertility (which they seemed to do in the Biblical case; consider how old most of the patriarchs were when they had their first children).
Again, if you think about it, the first humans were hunter-gatherers. Farming and pastoralism came later -- exactly as happened in the Bible. In the world of science, the New Stone Age began ten thousand years ago, in the world of the Bible, six thousand years ago. A trivial difference, in geologic terms; the Bible by this token appears to have preserved a folk memory that was largely forgotten elsewhere.
Add it all up, and it seems to me that the fact that Genesis so closely parallels the conclusions of scientists is evidence for the truth of Genesis -- it's being independently verified, even if there are a few footnotes being added.
The real stumbling block, it seems to me, is the word "day." The Big Bang (or the Steady State, or any other scientific model of the universe) didn't all happen in 144 hours -- the first part took moments, and the rest took billions of years.
But what is a day? On earth, now, it is 24 hours. It was somewhat less in the past -- the day was a few seconds shorter even in Biblical times; tidal effects are slowing earth's rotation. Nor is it a constant slowing; sometimes, for peculiar reasons, the earth's rotation suddenly changes ever so slightly.
Now consider this: On Jupiter, a day is about twelve hours.
On Venus, a day is 243 earth days -- longer than Venus's period of rotation around the sun (that is, a Venus day is longer than a Venus year).
An object in the Kuiper Belt beyond Neptune, if it is phase locked with the sun (as is likely true in many cases), will have a day in excess of 100 earth years. An object in the Oort Cloud, if it is phase locked, will have a day of thousands of years.
Theoretically, there could be a rock orbiting the sun with a year, and hence a phase-locked day, of two billion years (which would make seven days the total age of the universe so far). Kepler's third law tells us that this planet would be at a distance of a bit under 1,600,000 A.U. (where one A.U. or Astronomical Unit is the distance from the Sun to the Earth). That works out to a distance of about 150,000,000,000,000 miles, or 240,000,000,000,000 kilometers. That's a distance of 25 light-years, which means that that planet does not in fact exist in our solar system -- but, conceivably, there might be such a planet around an isolated star somewhere between galaxies. A planet with a period of half a billion years would be only 625,000 A. U. away; a planet with a period of a hundred million years would be 200,000 A. U. away. It is thus genuinely possible that there is a planet with a day of about 25 million years orbiting the sun.
The sun orbits the center of the Milky Way in a period estimated at 220 million years. The sun has a separate day, but a planet at this distance might be phase locked to the center of the galaxy and so have a 220 million year long day. The outermost stars of the galaxy appear to have an orbital period of over half a billion years. The data I have on this is sketchy, but it is clear that a planet orbiting the Milky Way at a sufficient distance, if it is phase locked, could have a day of two billion years.
On the other hand, an extrasolar planet was discovered in 2005 that has a year that is less than two earth days long. If there are any planets in that solar system even closer to the sun, they might have a year that is less than a day long!
Even on Earth, a day is six months long at the north and south poles. Away from the poles, different places have daylight at different times. If, as some have argued, it was sunset (or high noon, or sunrise) at Jerusalem when the first light came, then the first day over the Pacific Ocean was not evening and morning but morning and evening.
And then there is relativity. Time dilation. For a person or object moving at a speed near the velocity of light, a subjective 24-hour period could be a hundred years, or a thousand, or even two billion.
If you went to a planet around Alpha Centauri or Epsilon Eridani, and it had intelligent inhabitants, and you were translating the Bible, how would you translate Genesis? "And there was evening, and there was morning, the first 86400 seconds, where one second is defined as 9192631770 periods of the light emitted by cesium-133 as it shifts electrons between the two lowest states"? Go ahead, try it in English and see how many converts you get.
So what is a day? Answer: Unless otherwise specified, it's almost any period of time. Classical Hebrew doesn't have words for "geological epoch," or "plasma," or "strong nuclear force." (It does have a word for dust, and -- interestingly in light of Genesis 3:19 -- we are made of the interstellar dust which gave rise to stars and planets.) If you're going to tell the story of the Big Bang in classical Hebrew, what can you do except use a word like "day" for "epoch" and "water" (i.e. liquid, fluid) for "plasma"?
And are we really so arrogant as to think that God runs the entire universe based on our local earthly timescale? A God great enough to create on this scale is surely not parochial; why create a revelation that works only on one planet?
The fact that early Biblical commentators interpreted "day" to mean "24 hours" is surely not binding on God! After all, a thousand years are to God as one day....